Section 1. Gamétogenèse
Processus communs aux gamètes des deux sexes
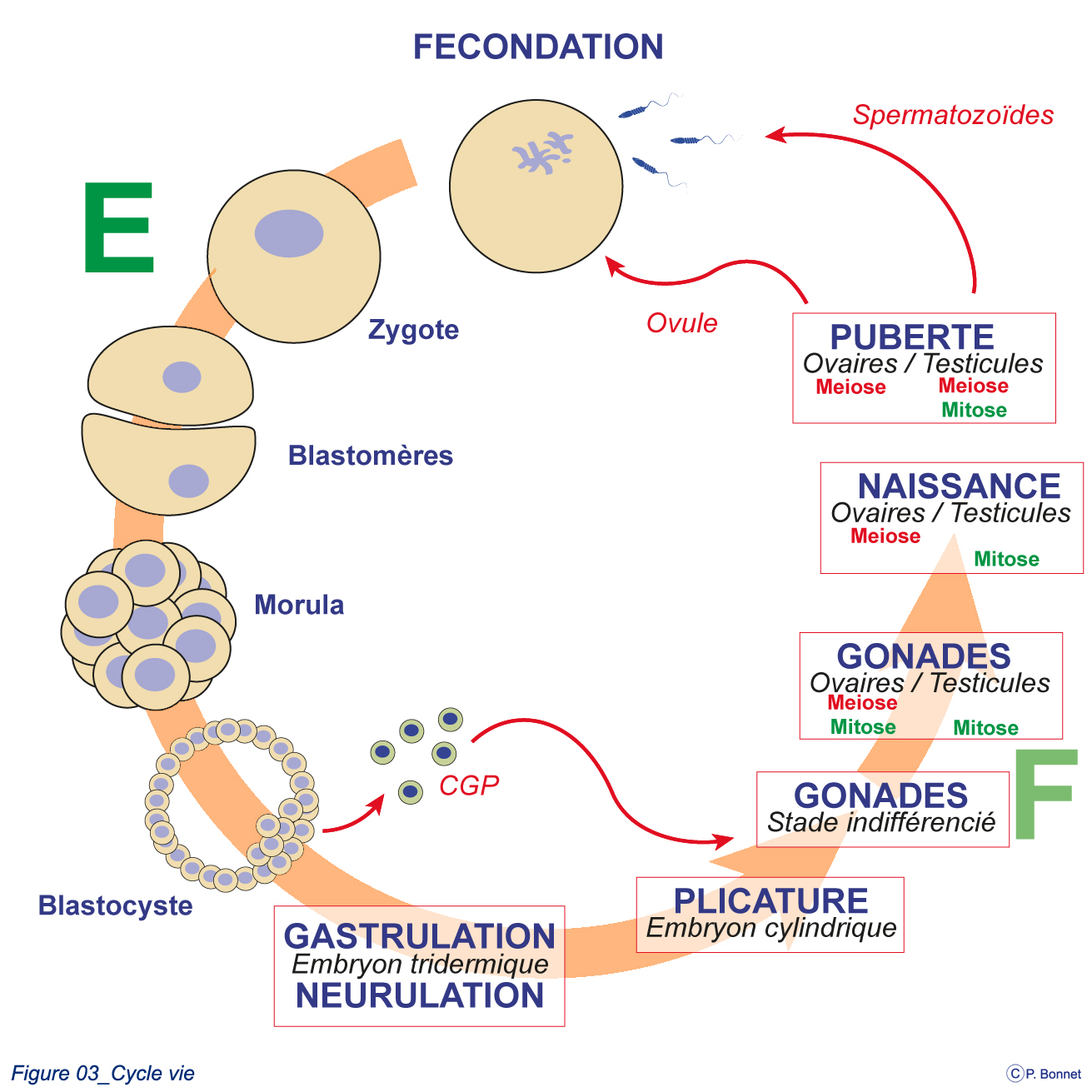
La formation des gamètes est initiée dès de début de la vie embryonnaire (figure 3-1).
|
|
|
|
Figure 3-1 |
Cycle de formation des gamètes |
|
Le zygote formé par la réunion des deux gamètes se divise pour donner les différentes formes successives de l’embryon (E), au stade du blastocyste, les cellules germinales primitives s’individualisent (CGP) et se placent à proximité du sac vitellin. Elles incorporeront en second lieu l’embryon pour se localiser dans la gonade primitive indifférenciée qui évoluera en ovaire ou testicule lors de la période fœtale (F). La division des CGP se fait par mitose puis par méiose suivant un « calendrier » différent chez la fille ou le garçon, pour finalement donner les gamètes mâles ou femelles. |
|
Les cellules destinées à la formation des gamètes s’individualisent de l’épiblaste (cfr. sections 3 et 4 de ce chapitre) lors de la troisième semaine de la vie embryonnaire et constituent les cellules germinales primordiales (CGP). Les CGP, douées de mouvements amiboïdes migrent hors de l’embryon pour se localiser dans la paroi de la vésicule vitelline près de l’allantoïde (voir figure 3-2 et la définition de ces structures dans les sections suivantes de ce chapitre).
Aidées par la flexion de l’embryon entre la quatrième et la sixième semaine, elles suivent le canal vitellin et l’intestin pour rejoindre les régions rétro-cœlomiques droite et gauche, à proximité des crêtes urinaires (figure 3-2). Celles-ci contiennent le rein intermédiaire (mésonéphros ou appareil de Wolff) (voir la mise en place des systèmes et organes dans la section 5 de ce chapitre ainsi que le chapitre traitant des systèmes uro-génitaux).
|
|
|
|
Figure 3-2 |
Migration des CGP |
|
A : Plan de section transversal T d’un fœtus en fin de plicature, le tube digestif est vu par transparence (jaune), Ce = céphalique, Do = dorsal et Ca = caudal. B : Portion caudale de la section : 1 = crêtes urogénitales, 2 = CGP migrant le long dui canal vitellin, 3 = sac vitellin, 4 = cavité coelomique, 5 = tube neural, 6 = ébauche vertébrale. |
|
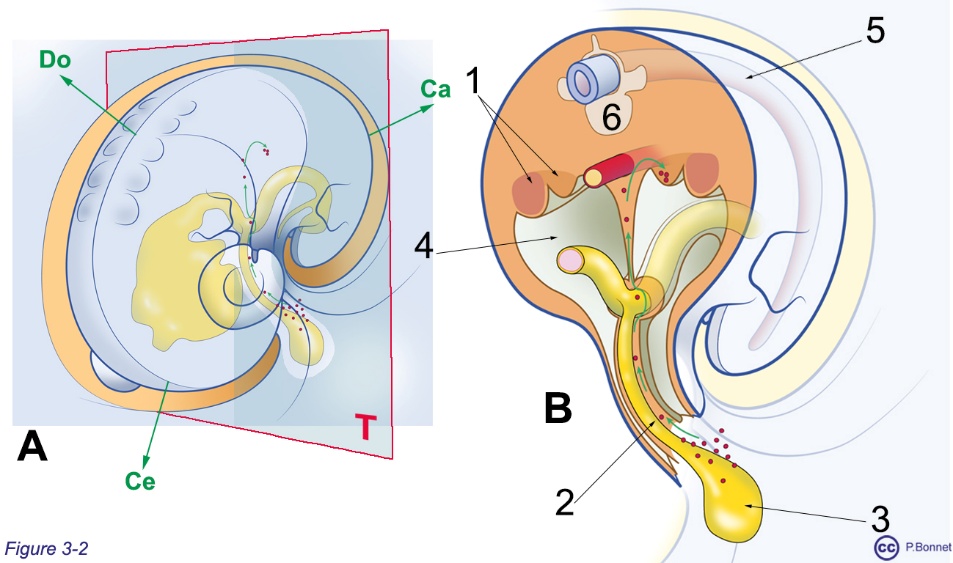
L’épithélium cœlomique s’épaissit en regard des CGP pour former les crêtes uro-génitales, où se développe le système paramésonéphrotique ou appareil de Müller (voir aussi le chapitre 10, section 1, figures 10-4-6-7-9-10-13 et 14).
Les CGP sont diploïdes et se multiplient par mitose lors de leur séjour extra-embryonnaire, de leur migration et au sein des crêtes urogénitales.
Les cellules coœlomiques situées en regard des crêtes urogénitales vont former des cordons vers la profondeur. Elles englobent les CGP et constituent les cordons sexuels , c’est l’ébauche gonadique indifférenciée (figure 3-3).
|
|
|
|
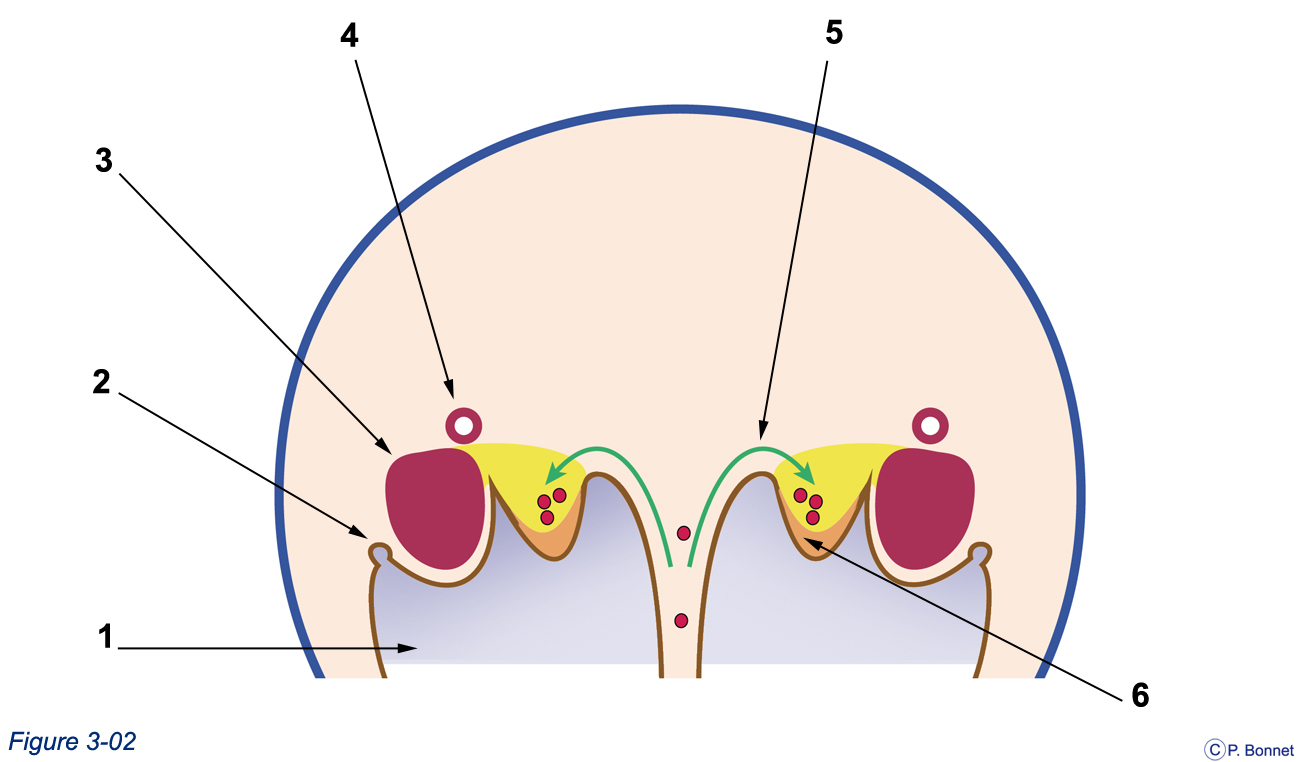
Figure 3-3 |
Gonade indifférenciée |
|
Coupe simplifiée correspondant à la section T de la figure 3-2, tube neural et ébauche vertébrale non représentés. 1 = Cavité cœlomique tapissée de l’épithélium cœlomique (brun), 2 = formation du canal paramésonéphrotique (de Müller), rein intermédiaire (mésonéphros), 4 = système collecteur du rein intermédiaire ou canal mésonéphrotique (de Wolff), 5 = trajet des CGP, 6 = épaississement de l’épithélium cœlomique donnant la crête génitale. |
|
Différenciation mâle / femelle
C’est la présence du gène SRY (Sex determining Region of the Y chromosome) qui déterminera l’évolution vers une gonade mâle (testicule). En l’absence de ce gène, l’évolution se fera vers une gonade femelle (ovaire) (figure 3-4).
|
|
|
|
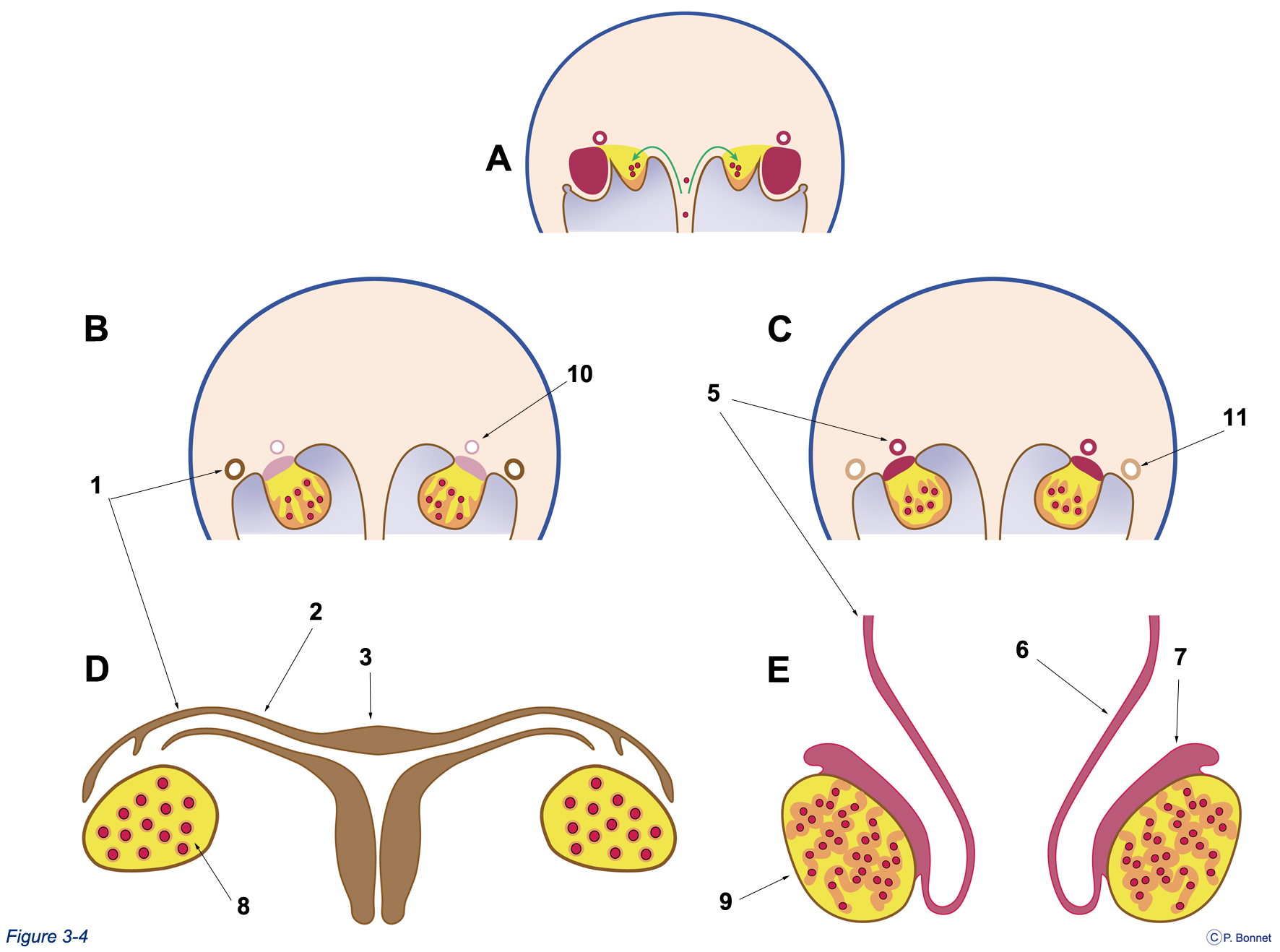
Figure 3-4 |
Formation de l’ovaire et du testicule |
|
A, B et C : Coupes similaires à la coupe de la figure 3-3. D : Coupe simplifiée des organes génitaux internes femelles. E : Coupe simplifiée des organes génitaux internes mâles. À partir du stade indiférencié (A), en l’absence du gène SRY, le système Müllerien persiste (1) pour donner, entre autres, la trompe (2) et l’utérus (3). Le système Wolffien régresse (10), la gonade est l’ovaire (8). Si le gène SRY est présent, le système Mullérien disparaît (11), le système Wolffien persiste (5) pour donner, entre autres, le canal déférent (6) et l’épididyme (7), la gonade est le testicule (9). |
|
Formation de la gonade mâle et spermatogenèse
Sous l’influence du gène SRY (chromosome Y) exprimé au sein des cellules de la crête génitale, les cordons sexuels prolifèrent et deviennent des cordons pleins, des cordons séminifères des ou cordons testiculaires.
À la puberté, ces cordons se creusent pour former des tubes, les CGP qu’ils contiennent deviennent alors des spermatogonies (cellules germinales primordiales différenciées).
À partir de la puberté, les spermatogonies (de type A) se divisent par mitose, donnant une spermatogonie (de type B) qui rentre dans le processus de spermatogenèse et une autre qui renouvelle le stock.
La spermatogonie de type B se divise par plusieurs mitoses pour aboutir aux spermatocytes de type I.
Les spermatocytes de type I s’engagent dans la première division méiotique pour donner les spermatocytes de type II qui deviendront les spermatides à la suite de la deuxième division méiotique.
Ces étapes de la spermatogenèse constituent la spermacytogenèse.
Les spermatides seront ensuite maturées en spermatozoïdes, cette deuxième étape de la spermatogenèse constitue la spermiogenèse. Lors de ce processus, le noyau se condense, le flagelle et l’acrosome apparaissent (figure 3-5).
|
|
|
|
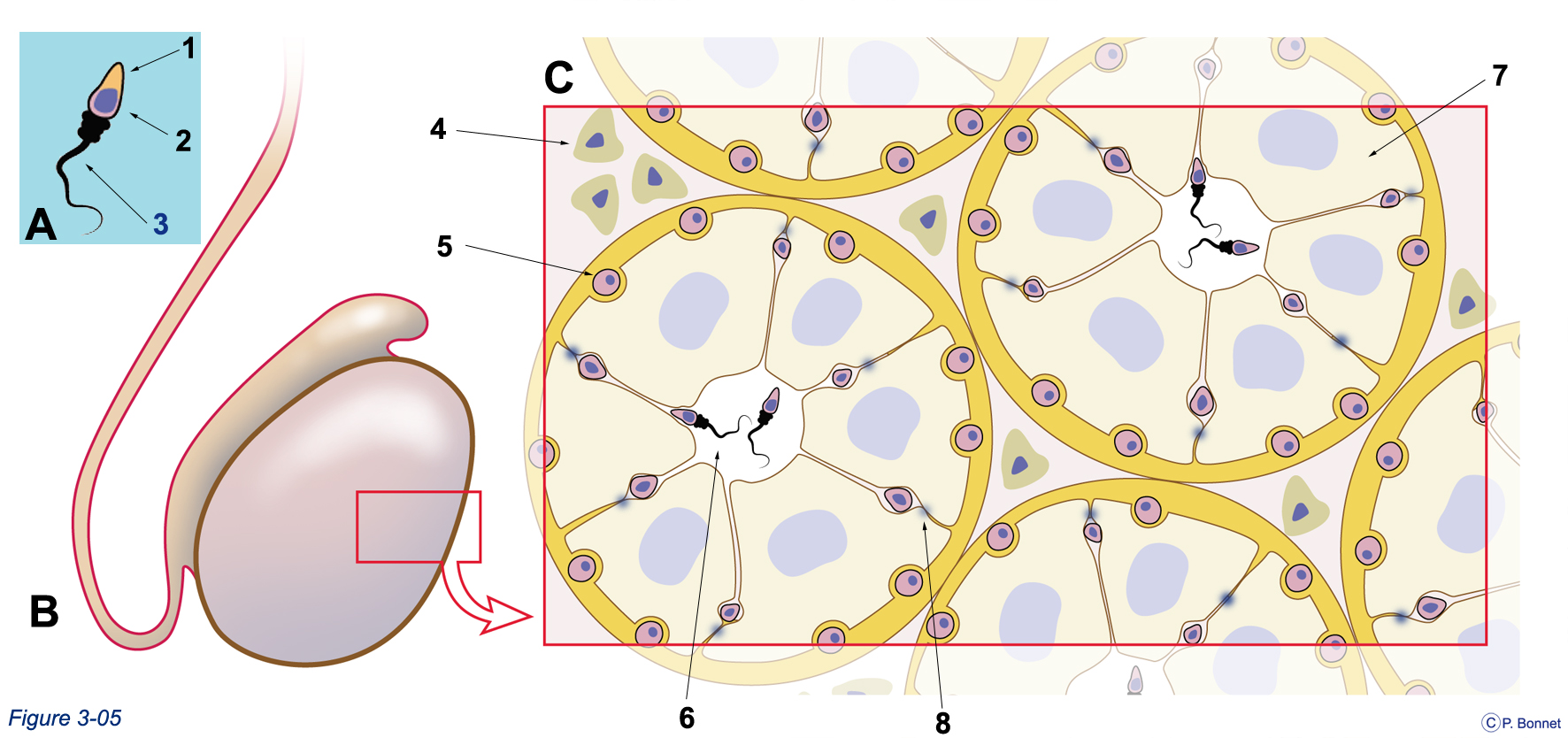
Figure 3-5 |
Gonade mâle et spermatogenèse |
|
A : Spermatozoïde formé, 1 = acrososome, 2 = noyau condensé, 3 = flagelle et pièce intermédiaire. B : Testicule dans lequel une portion (rectangle rouge) est prélevée pour examen microscopique. C : Schéma simplifié de l’observation microscopique du parenchyme testiculaire. Entre les tubes séminifères coupés transversalement et délimités par de grandes cellules (Sertoli) = 7, se situent les cellules de Leydig = 4, secrétant la testostérone. Les spermatogonies = 5 se multiplient par mitose en périphérie des tubes ou s’engagent par méiose dans le processus de formation des spermatozoïdes. Celui-ci a lieu entre les cellules de Sertoli de la périphérie au centre du tube pour aboutir à la formation d’un spermatozoïde dans la lumière du tube. La lumière du tube communique avec l’épididyme pour arriver aux voies séminales. Rapidement, les spermatogonies s’engageant dans le processus méiotique et localisées entre les cellules de Sertoli, sont isolées des cellules périphériques et du compartiment sanguin par des jonctions serrées (= 8) entre les faces latérales des cellules de Sertoli. Il existe ainsi une barrière hémato-testiculaire. |
|
Formation de la gonade femelle et ovogenèse
En l’absence du gène SRY, les cordons sexuels prolifèrent tout en gardant un contact avec l’épithélium cœlomique. Les CGP se multiplient par mitose de la 15e semaine au 7e mois pour donner des ovogonies. Progressivement ces ovogonies entre dans le processus de division méiotique et portent alors le nom d’ovocytes primaires, les cellules des cordons s’organisent au pourtour des ovocytes primaires et forment les follicules primordiaux. À la naissance, les ovocytes primaires restent bloqués au niveau de la première division méiotique et le stock d’ovocytes est donc définitivement constitué
L’évolution du follicule primordial va se poursuivre à deux niveaux (figure 3-6) :
- Au niveau de l’enveloppe cellulaire, l’évolution sera surtout importante lors de la puberté.
- Au niveau de l’ovocyte primaire le processus de division méiotique redémarre exclusivement à la puberté.
Le follicule primordial évolue suivant les étapes suivantes :
- Les cellules entourant le follicule deviennent cubiques, on parle alors de follicule primaire.
- Lorsque la couche des cellules entourant l’ovocyte devient pluricellulaire, on atteint le stade du follicule secondaire. Une couche de glycoprotéines se constitue entre ces cellules et l’ovocyte : la membrane pellucide. Un bon nombre de follicules primaires involuent cependant.
- Si le follicule continue d’évoluer, il se transforme en follicule tertiaire : une cavité se forme au sein de l’amas de cellule entourant l’ovocyte. Les cellules du stroma ovarien s’organisent en thèques interne et externe.
- À la moitié de la première phase du cycle menstruel, un ovocyte tertiaire sur les deux ovaires devient dominant et croît pour devenir le follicule de de Graaf de 25 mm de diamètre.
|
|
|
|
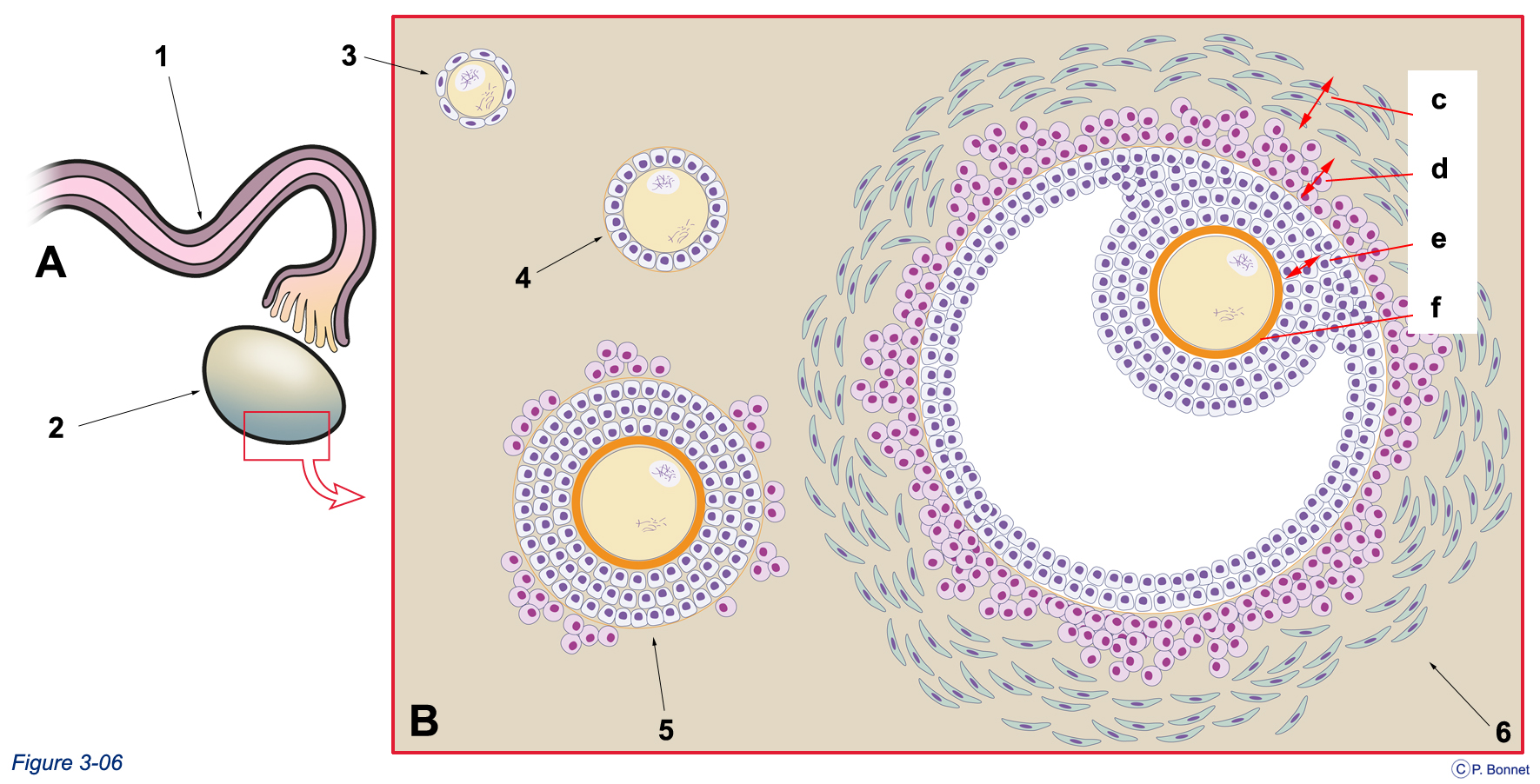
Figure 3-6 |
Gonade femelle et formation des follicules |
|
A : Ovaire (2) surmonté de la trompe (1), aspect macroscopique. Le rectangle rouge indique la zone prélevée pour analyse microscopique. B : Schéma simplifié de l’organisation des différents types de follicules au sein du parenchyme ovarien. 3 = follicule primordial entouré des cellules coelomiques, 4 = follicule primaire (cellules cubiques), 5 = follicule secondaire avec sa membrane pellucide (orange), 6 = follicule tertiaire (cavitaire) pouvant évoluer vers un follicule de de Graaf (c = thèque externe, d = thèque interne, e = corona radiata, f = membrane pellucide). |
|
Au sein du follicule dominant, le pic sanguin de LH (Hormone Lutéinisante) qui marque le milieu du cycle menstruel provoque la fin de la première division méiotique de l’ovocyte primaire. L’ovocyte primaire (deux jeux de chromosomes redupliqués) donne ainsi deux cellules filles : l’ovocyte secondaire et son premier globule polaire qui contiennent chacun un jeu de chromosomes redupliqués. La deuxième division méiotique débute et s’arrête en métaphase.
Le follicule évolue également : l’ovocyte secondaire entouré d’une couronne de cellules (la corona radiata) et de sa membrane pellucide se détache de la paroi folliculaire.
38h00 après le pic de LH, le follicule se rompt et libère le liquide folliculaire et l’ovocyte qui seront récupérés par la trompe utérine : c’est l’ovulation. La deuxième division méiotique sera achevée lors de la pénétration du spermatozoïde :
- L’ovocyte secondaire donnera deux cellules : l’ovocyte mature (qui sera le seul à fusionner avec le spermatozoïde) et le deuxième globulaire.
- Le premier globule polaire donnera deux globules polaires.
|
||||||||||||||||||||||
|
Figure 3-7 |
Tableau comparatif de la formation des gamètes mâle et femelle |
|||||||||||||||||||||
